Eli Broemer
/E·Lye Bray·mer/
 Home
HomeAbstract
Full PDFInflammasome NLRC4 Gene Knockout Increases Stiffness of Bladder Wall in Rat Model
Eli Broemer (1), Christina Chan (2), Sara Roccabianca (1)
(1) Mechanical Engineering
Michigan State University
East Lansing, MI, USA
(2) Chemical Engineering
Michigan State University
East Lansing, MI, USA
INTRODUCTION
The urinary bladder is a highly compliant organ with its mechanical behavior largely mediated by smooth muscle and the extracellular matrix (ECM) in the bladder wall [1]. The ECM is susceptible to pathological remodeling wherein the structure of its constituents—collagen and elastin—is altered which impacts stiffness and bladder health/function. Loss of healthy bladder function can reduce the affected individual’s quality of life: increasing the risk for depression and suicide, or unhealthy coping methods such as reducing fluids and isolation [2]. Annual healthcare cost for treating poor bladder function in the United States is estimated between $9-100 billion [2, 3].
Common urinary dysfunctions have been associated with an inflammatory response including: neurogenic bladder, chronic urinary tract infection, and interstitial cystitis (or, bladder pain syndrome) [4]. Furthermore, inflammation is believed to be involved in the initiation and progression of these dysfunctions. Chronic inflammation within the bladder wall is associated with ECM remodeling which can damage the organ tissue in the long term [5].
Inhibiting pro-inflammatory mechanisms may be a viable approach to prevent bladder wall remodeling and function deterioration. Specifically, inflammasome- mediated pathways has emerged as driving fibrosis in various organs including the bladder [6]. For example, the NOD-like receptor (NLR) inflammasomes NLRP3 and NLRC4 (Pyrin domain-containing protein 3, and Caspase recruitment domain- containing protein 4, respectively) initiate inflammation in response to environmental factors, including gram-negative bacteria such as E. coli [7]. NLRC4 is found throughout the human body, and in the urothelium—the protective layer between urine and bladder tissue—of the bladder wall [8].
Knockout (KO) of the NLRC4 gene may reduce bladder inflammation as it is functional in activating immune response in the bladder wall [8]. In the first stage of this study, we show that NLRC4 KO alone affects mechanical behavior of bladder ECM in rat models.
METHODS
An NLRC4 KO rat model was developed at the MSU Transgenic and Genome Editing Facility by deleting 252 base pairs corresponding to the NLRC4 gene in Sprague- Dawley (SD) rats. Then heterozygous (NLRC4 +/-) and homozygous (NLRC4 -/-) groups were bred. Controls were SD wild-type. Animals were sacrificed at 12-13 weeks, and whole bladders were extracted, imaged, weighed, sectioned into rings, and frozen until testing. Dissection and testing was randomized and blinded.
Fresh tissue geometry was assessed from top-down photos of the flattened whole organ, as shown in Figure 1a. Bladder area was measured by thresholding these profile shots with ImageJ, as shown in Figure 1b. As the size and weight of the extracted organ is somewhat dependent upon the location of incision at the bladder neck, these measures were normalized by ratio of bladder weight to area.
After thawing, bladder ring samples were decellularized to isolate the ECM, and mounted to a uniaxial testing machine while submerged in Krebs solution. Tests measured force via load cell and length/cross-sectional area via front/side cameras.
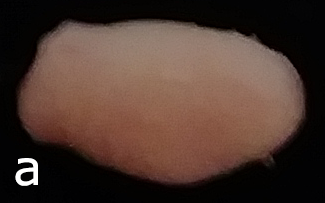

Figure 1: a) Whole fresh bladder after extraction flattened to show profile, b) Threshold applied to measure bladder profile area.
Samples were stretched to four strain levels (5, 10, 15, and 20%). For each of the four tests, the tissue was held at a 2g preload until relaxation diminished, and then cyclically loaded and unloaded at 1% strain/sec. The first test had 10 minute preload and 10 cycles, while the next three tests had 1 minute preload and 5 cycles. This loading protocol ensures each ring is consistently taut and preconditioned so that mechanical behavior becomes repeatable.
Data from the final load cycle at the highest strain level was analyzed and is presented in Figure 2. True (Cauchy) stress-strain curves were found using the median cross-sectional area at the preload stage, approximated as a rectangle, and then force and length, recorded continuously, give stress and strain assuming incompressibility. The initial length (reference) used to evaluate stress and strain was each sample’s length at the start of the last loading cycle.
RESULTS
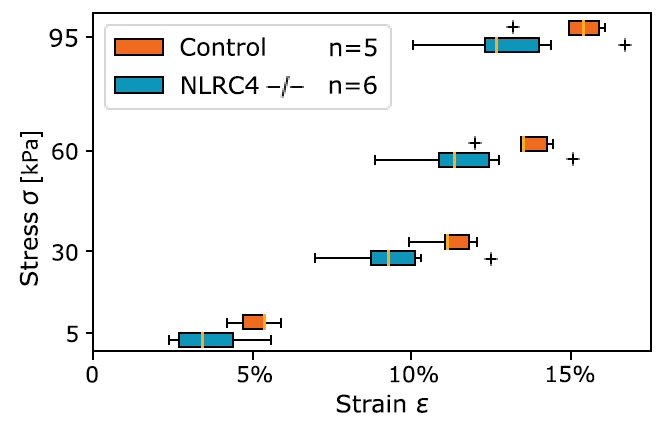
Figure 2: Cauchy stress-strain curve at representative stress levels from bladder ECM tests. Student’s t-test of Control v NLRC4 -/- gives P=.049, P=.09, P=.11, and P=.11 at 5, 30, 60, and 95 kPa respectively (no correction).
Figure 2 shows the circumferential mechanical behavior, tested uniaxially, for preconditioned tissue stretched to 20% strain. It should be noted this data is truncated to ~15% strain to better show its non-linear characteristic. The bladder ECM for the KO group (NLRC4 -/-) shows less deformable mechanical characteristics when compared to controls. This difference between control and KO groups was significant at lower values of stress (i.e., 5 and 30 kPa). At higher values of stress (i.e., 60 and 95 kPa), the trend is conserved yet the difference is not significant (P=.11 for both stress values). Heterozygous (NLRC4 +/-) had values of strain in-between control and KO groups for all stress values considered (differences were not significant when compared to either the control or KO animals).
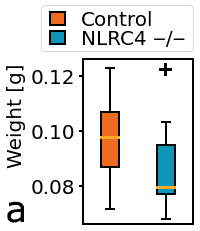
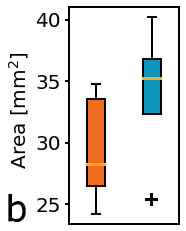
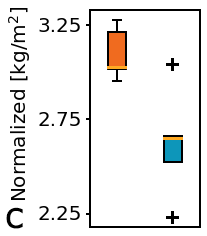
Figure 3: Physical characteristics of fresh whole bladders; a) Weight, b) Flattened area, c) Normalization by weight/area.
Figure 3a shows that whole bladder weight was lower in the KO group compared to controls. Bladder profile area appeared larger in the KO group compared to controls (Figure 3b). Normalization of fresh bladder weight and flattened area (Figure 3c) showed marked difference between KO and control groups.
DISCUSSION
These results indicate that NLRC4 KO affects the homeostatic mechanical characteristics of the urinary bladder’s ECM. Interestingly, while high distensibility is often problematic and a clinical marker for bladder dysfunction, bladders having comparatively lower distensibility are considered to be stable and healthy [9, 10]. Our data show that NLRC4 KO results in a less distensible ECM and in a lower bladder weight, when compared to controls. This has also been observed in children, where low bladder weight, assessed by ultrasound, was used to predict lower distensibility, as compared to dysfunctional highly distensible bladders [9].
Bladder weight is also considered a marker for bladder inflammation, indeed both NLRC4-driven and bacterial-induced inflammation have shown to double bladder’s weight compared to controls [11, 12]. Moreover, bacterial-induced inflammation can trigger detrusor hyperactivity [12], which is compensated for by increased bladder distensibility [13]. Taken together, previously published literature and the results presented here, suggests that the NLRC4 -/- animals had lower levels of inflammation than the control.
Increased bladder area was observed in the KO group vs controls, although reference length (a measure of ECM ring circumference) was similar across groups being 16.8mm +/- 0.5mm. This apparent size discrepancy may be clarified by noting area was measured in relaxed bladders having muscle cells, while reference length was measured in decellularized preconditioned tissue. The difference revealed by normalization may be due to microstructural changes in the detrusor muscle and/or ECM, though that is yet to be determined by histology.
Next steps in this study include histological and immunohistological studies to examine potential microstructural changes between groups as well as the presence and level of inflammatory markers, such interlukin-1β (IL-1β), IL-18 and IL-6. Later steps will include the addition of inflammatory stressors such as a high-fat diet or diabetic predisposition.
ACKNOWLEDGEMENTS
We thank Drs. Elena Demireva and Huirong Xie from the MSU Transgenic and Genome Editing Facility and the Institute of Quantitative Health Science and Engineering for developing the NLRC4 KO rat model. We thank fellow lab members Marissa R. Grobbel and Tyler Tuttle for assistance with mechanical testing protocol. This study was supported by NSF grant CBET 1802992.
REFERENCES
[1] Parekh, A et al., J Biomech, 43(9):1708-16, 2010. [2] Sacco, E et al., J Urol, 2:11-24, 2010. [3] Powell, LC et al., Neurourol Urodyn, 37(4):1241-49, 2018. [4] Liu, HT et al., Low Urin Tract Symptoms, 7(2):102-7, 2015. [5] Kullmann, FA et al., Front Syst Neurosci, 12:13, 2018. [6] Inouye, BM et al., Curr Urol, 11:57-72, 2017. [7] Zheng, D et al., Cell Discovery, 6:36, 2020. [8] Hughes Jr., FM et al., Int Urol Nephrol, 47(12):1953-64, 2015. [9] Ukimura, O et al., J Urology, 160(4):1459-62, 1998. [10] Bauer, SB, Pediatr Nephrol, 23:541-51, 2008. [11] Hughes Jr, FM et al., Am J Physiol-Renal, 306(3):F299-308, 2014. [12] Randich, A et al., J Pain, 7(7):469-79, 2006. [13] Howard, PS, Macarak, EJ, Adv Exp Med, 462:215-223, 1999.
SB3C2021
Summer Biomechanics, Bioengineering, and Biotransport Conference
June 14-18, Virtual